染色体的结构变化主要包括哪些内容
编辑推荐:
(2010-08-27)
- (2010-08-27)
- (2010-08-27)
- (2010-08-27)
染色体结构畸变 染色体的结构变化主要有以下几种:
①缺失 染色体臂发生断裂并丢失一部分遗传物质的结果。一个染色体臂发生了断裂,而这种断裂端未能与别的断裂端重接,那么就形成一个带有着丝粒的片段和一个没有着丝粒的片段。后者在细胞分裂过程中不能定向而被丢失。带有着丝粒的片段便成为一个发生了末端缺失的染色体。如果一个染色体发生两次断裂而丢失了中间不带有着丝粒的片段,留下的两个片段重接以后便成为发生了中间缺失的染色体。如果同一染色体的两臂同时发生了断裂,而余下的两臂的断面间又发生重接,便形成环形染色体。根据所丢失的染色体片段的大小,缺失所带来的危害性也各不相同。较大的缺失往往带来致死效应,而微小的缺失则并不致死。如果缺失部分包括某些显性等位基因,那么同源染色体上与这一缺失相对应位置上的隐性等位基因就得以表现,这一现象称为假显性。在玉米中,如果染色体上带有颜色决定基因的区段缺失则常能产生特定的表型效应,例如白苗与褐色中脉等。在人类中,染色体的部分缺失常导致染色体病,如猫叫综合征就是由于5号染色体的短臂部分缺失所致。
②重复 一个染色体上某一部分出现两份或两份以上的现象。首尾相接的重复称为衔接重复或串接重复;首尾反方向连接的重复称为颠倒衔接重复或倒重复。重复部分可以出现在同一染色体上的邻近位置,也可以出现在同一染色体的其他位置或者出现在其他染色体上。重复杂合体具有特征性的减数分裂图象,它的染色体在进行联会时重复片段在同源染色体上找不到相应的结构,因而形成称为重复环的环状突起。类似的图象可以在果蝇的重复杂合体的唾腺染色体中看到。在缺失杂合体细胞中也同样可以看到图象相似的缺失环。重复的遗传效应比缺失来得缓和,但重复太大也会影响个体的生活力,甚至引起个体的死亡。染色体上某些区域的重复可以产生特定的表型效应,例如果蝇的显性基因棒眼(Bar eye,B)就是重复的结果。主要的表型效应是复眼中的单眼数减少,使复眼呈棒状而不是通常的卵圆形。在这种果蝇的唾腺染色体上可以看到X染色体上明显的横纹重复(见位置效应)。可是对于一般的染色体来说,不通过显带法是很难检出重复的。
③倒位 一个染色体上同时出现两处断裂,中间的片断扭转180°,重新连接起来而使
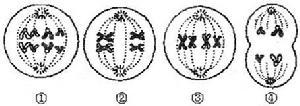
这一片段的基因的排列顺序颠倒的现象。颠倒片段包括着丝粒的倒位称为臂间倒位;不包括着丝粒的倒位称为臂内倒位。两个断裂点与着丝粒之间的距离不等的臂间倒位是容易识别的,等距离的倒位则除非应用显带技术一般不易察觉。倒位杂合体也具有特征性的减数分裂图象,它的倒位染色体和正常同源染色体联会时出现倒位环。。臂内倒位杂合体如果在倒位环内发生一次交换便形成一个有两个着丝粒的染色单体和一个没有着丝粒的片断,这样在减数分裂后期就会出现染色体桥和没有着丝粒的片段,后者往往无法进入子细胞核中;而有两个着丝粒的桥被拉断后,虽然两个染色体可分别进入子细胞,但由于断裂位置不一,往往带来缺失而导致配子的死亡。
④易位 一个染色体臂的一段移接到另一非同源染色体的臂上的结构畸变。两个非同源染色体间相互交换染色体片段称为相互易位。相互易位的染色体片段可以是等长的,也可以是不等长的。一般基因改变它在染色体上的位置时并不改变它的功能,可是在果蝇等生物中发现如果位置在常染色体的基因通过易位而处于异染色质近旁时,它的功能便会受到影响而呈现花斑位置效应现象。倒位也可能带来同一效应。易位纯合体没有明显的细胞学特征,它在减数分裂时的配对也不会出现异常,因而易位染色体可以从一个细胞世代传到另一个细胞世代。可是易位杂合体则不同,由于正常染色体和易位染色体的同源部分的配对,在减数分裂中期可以看到特有的十字形图象。
随着分裂过程的进行,十字形逐渐开放,邻接的两个着丝粒趋向同一极或趋向两极,形成环形或8字形图象。前一种染色体离开方式称为邻近离开,后一种方式称为交互离开。相互易位杂合体的花粉母细胞中大约有50%的图象呈环形,属邻近离开,50%呈8字形,属交互离开。这说明四个着丝粒趋向两极的取向是随机的,行动是***的。邻近离开的结果使配子内含有重复或缺失的染色体,形成致死的不平衡配子。交互离开导致非致死的平衡配子,其中半数配子的染色体是正常的,半数配子具有平衡的易位染色体,这就是说虽然发
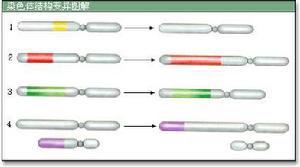
生了易位,可是易位并不带来基因的增加或缺少。交互离开使两个易位染色体进入一个配子细胞,两个非易位染色体进入另一配子细胞中。所以这种分离方式使非同源染色体上的基因间的自由组合受到限制,使原来在不同染色体上的基因出现连锁现象,这种现象称为假连锁(见基因定位)。
两个或两个以上染色体之间发生相互易位时,如果这些染色体的近心端相接,就形成双着丝粒体或多着丝粒体。双着丝粒体同时具有两个有功能的着丝粒,它们在细胞分裂后期各自趋向两极,从而出现后期桥。如果这种桥被拉断,往往导致细胞死亡。然而根据玉米胚乳细胞的研究,认为染色体桥虽可被拉断,但两端的两个着丝粒仍可顺利地进入细胞的两极并参与子细胞的形成,而且它们的断裂端仍然开放着并可重接。当再次相接后就会在下一次分裂的后期形成另一个桥,这种“断裂-融合-桥”周期可以持续许多个细胞世代。
专业考试题库一套,历年真题、章节
练习题、考试模拟题应有尽有!
手机/微信:17052762074

本网所注明来源为"爱爱医"的文章,版权归作者与本站共同所有,非经授权不得转载。
本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们
联系,我们将立即进行删除处理。
- [广告]在新利网官方网站 报培训班的考生赠送新利网官方网站 vip服务(班主任老师全程指导、量身制定学习计划、学习跟踪、学习督导、专家在线答疑等);赠送新利网官方网站 VIP题库1个。
- 课程名称
- 课程试听
- 报名
 免费试听
免费试听
- 推荐阅读
- · 收藏!医师资格考试报名材料清单汇总!

