黏着斑激酶在酒精性肝病中作用的研究进展
酒精性肝病(alcoholicliverdisease,ALD)是由于长期饮酒所致的肝脏疾病,疾病初期通常表现为脂肪肝,进而可发展成酒精性肝炎、酒精性肝纤维化和肝硬化。在我国,酒精性肝病的发病率呈逐年上升趋势,乙醇对肝脏有明显的毒性作用,重度饮酒者中80%以上有一定程度的脂肪肝,10%N35%可发展成酒精性肝炎,10%~20%将发展为肝硬化。但关于酒精性肝病的发病机制目前尚未完全阐明。大多数细胞,包括肝细胞在内,与细胞外基质(extracellularmatrix,ECM)的相互作用影响着细胞的生存、生长和功能等。适宜的肝细胞-ECM条件不仅能维持良好的细胞位置和形态,还能通过各种细胞-ECM的信号通路影响细胞增殖、分化、凋亡、基因表达、细胞极性和基质合成等。肝细胞主要是通过细胞表面的受体与ECM黏附,其中最重要的是整合素家族,而黏着斑激酶(focaladhesionkinase,FAK)在整合素介导的信号转导途径中起着关键作用,其活化后对多种类型细胞的生物学行为发生作用,如黏附、迁移、增殖、分化、凋亡、基因表达及细胞周期调控等叫。本文主要阐述FAK在酒精性肝病发病机制中的作用。
一、FAK的结构与活性
FAK基因位于人染色体的8q24,全长4285bp,能够编码1052个氨基酸。蛋白结构是一种非受体胞质酪氨酸激酶,相对分子质量为125×l03。1992年,FAK由Hanks等从v-src转染的鸡胚成纤维细胞中发现,果蝇、斑马鱼、鸟、鼠、非洲蟾蜍、人类等多种生物中均有表达,且有高达90%的同源性。
FAK蛋白质结构由可分为3个功能区,即N-端区、激酶区和C-端区(图1)。每个区域约包含400个氨基酸。氨基酸390~650位是中间的激酶区,就是高度保守的催化区域,两端分别为氨基端和羧基端脚。N-端区包括FERM同源区,缺乏N-端区的FAK分子(FAK△384)在受到**时的磷酸化水平明显高于野生型的FAK分子,可见FERM区可抑制FAK活性,外界**能解除这种抑制作用,而使FAK发生活化。C-端区包含黏着斑靶向序列(focaladhesiontargeting,FAT),可结合paxillin和talin两个黏附相关蛋白协助FAK固定位于黏着斑。FAK相关非激酶(FAK-relatednon-kinase,FRNK)是C-末端基因单独表达的蛋白质,因包含FAT,可与FAK竞争,是FAK的内源性抑制剂吲。C-端区还含有两个脯氨酸富集区,是衔接蛋白p130cas与鸟苷三磷酸酶调节蛋白GRAF/ASAP的结合区,能发挥信号转导功能。
FAK蛋白包含6个酪氨酸磷酸磷酸化位点:Tyr397,Tyr407,Tyr576,Tyr577,Tyr861与Tyr925。其中Tyr397是最重要的自主磷酸化位点,其磷酸化能增强FAK的催化活性,并能与多种信号蛋白发生作用,如Src酪氨酸激酶家族、磷脂酰肌醇3-激酶(phosphatidylinosit013-kinase,PI3K)、生长因子受体结合蛋白7(growthfactorreceptor-boundprotein7,Grb7)等。当Src被募集活化后,能依次激活Tyr576,Tyr577,Tyr861与Tyr925磷酸化位点,而Tyr925磷酸化后能募集Grb2等接头蛋白。此外FAK能募集和激活多种信号蛋白,影响下游各种信号通路。
二、FAK在ALD中的作用
1.乙醇与FAK:French等通过动物实验,提出不同血液乙醇水平可以显著改变基因表达强度,蛋白水平和蛋白磷酸化水平。Bardag-Gorce等建立乙醇饮食大鼠模型,保持血液乙醇高浓度,约为100mmol/L,研究在慢性乙醇作用下各种信号通路基因表达的改变,其中发现黏着斑基因在长期乙醇喂养组的大鼠明显比对照组大鼠高表达。Tuma等恻也通过实验证明慢性乙醇饮食可导致细胞黏附过程的异常,这种损害在肝静脉周围肝细胞较门静脉周围的肝细胞明显,实验发现大鼠肝静脉周围的肝细胞中整合素p1含量明显升高,但门静脉周围的肝细胞整合素pl与控制组没有明显不同,并且提出这种肝细胞-ECM相互作用的改变会影响肝细胞的结构和功能,这一机制在乙醇造成的肝脏损伤中起作用。Schaffert等提出大鼠经2~3周的乙醇**后能增加肝静脉周围肝细胞中整合素p1、a1和a5亚基含量,但黏着斑蛋白中FAK、桩蛋白、黏蛋白水平没有明显改变,并提出慢性乙醇**可能改变了FAK的磷酸化水平,以此来调节黏着斑更新率,这一研究目前正在进行中。
目前多数学者认为FAK的活化依赖于整合素聚集成簇,整合素与基质蛋白的聚集或胞质区构象的改变可增强FAK的N-末端结构域与整合素D尾的结合,而整合素p的细胞质区是FAK活化所必需的,这种结合使FAK活化位点Tyr397暴露出来,并且自身磷酸化,继而导致Tyr576、Tyr577等相继磷酸化,最终激活整个酶区的FAK。整合素-ECM相互作用改变也主要是由于黏着斑的装配或者更新率发生变化。慢陛乙醇作用并不能改变肝细胞内FAK的分布,但可使肝细胞中黏着斑和肝静脉周围的整合素pl的含量增加,增加FAK磷酸化水平而使得其活性增强。
2.FAK与脂质代谢:FAK能够通过多种途径活化丝裂原活化蛋白激酶(mitogen-achvatedproteinkinases,MAPK)通路。FAK含有p130CAS接头蛋白的结合位点,p130CAS酪氨酸磷酸化能进一步聚集Crk、Nck接头蛋白与鸟嘌呤核苷酸交换因子(sonofsevenless,SOS)结合,从而**MAPK信号途径。FAK还含有Grb2结合位点,当Tyr925位点被磷酸化后能募集Grb2/SOS复合物形成。另外FAK的活化还可使Shc的酪氨酸磷酸化水平升高,促进Grb2/SOS复合物的形成,同样可以增强MAPK活化。由此激活Ras-Raf-丝裂原活化蛋白激酶激酶(mitogen-activatedproteinkinasekinase,MEK)-细胞外调节蛋白激酶(extracellularregulatedproteinkinases,ERK)通路,使下游的ERK表达增加。
近年研究发现ERK可使激素敏感脂肪酶(hormone-sensitivetriglayceridelipase,HSL)丝氨酸的第600位点磷酸化,HSL是脂解作用的关键酶之一,其活性增强可导致血清游离脂肪酸浓度升高,脂肪重新分布,引起脂肪肝等的发生。
3.FAK与枯否细胞:枯否细胞(Kupffercells,KC)是肝脏内的巨噬细胞,位于肝血窦内,与血浆直接接触,在乙醇诱导的肝脏损伤中具有重要作用,研究发现去除KC能够预防乙醇诱导的脂肪肝和肝脏炎症反应,从而减少了酒精性肝病的发生。
有研究显示慢性乙醇**可诱发ERKl/2的活化增强,激活转录因子Egr-l,使KC中的肿瘤坏死因子α(TNFα)mRNA水平增高,是对照组的2倍。而TNFα是脂肪肝重要的炎性因子之一,能激活一系列炎症反应,在脂肪肝的发病过程中起重要作用。慢性乙醇作用可使FAK磷酸化水平增加,并通过上述各种方式激活ERKl/2而发挥作用。
4.FAK与肝纤维化:酒精性肝病可表现为肝纤维化,多发生在肝细胞和静脉周围细胞,这也与实验中慢性乙醇饮食导致大鼠整合素p1含量升高在肝静脉周围尤其明显相一致。这可能是由于FAK在肝星状细胞(hepaticstelLateceU,HSC)的增殖及肝纤维化形成过程中发挥了重要作用。在大鼠胆总管结扎肝纤维化模型中,随着肝纤维化程度逐渐加重,肝组织FAK磷酸化水平有所增强。
FAK-PI3K-Akt信号通路在HSC增殖中发挥重要作用。Reif等研究发现抑制FAK活性可以阻止血小板衍生生长因子(platelet-derivedgrowthfactor,PDGF)诱导的PI3K和Akt活化,抑制HSC增殖;PI3K抑制剂LY294002、FAK和Akt的突变体均可抑制PDGF诱导的HSC增殖。
FAK磷酸化能调控细胞周期相关蛋白,促进HSC增殖,使肝纤维化形成。Zhao等应用细胞周期蛋白1(cyclinDl)启动子荧光酶分析,FAK能通过ERK通路在转录水平调节eyclinDl的表达,实验证明ERK特异性抑制剂PD98059可以抑制FAK活化,减弱cyclinDl的表达。p160ROCK的特异性抑制剂Y27632干预大鼠HSC后,发现FAK与ERK磷酸化受阻,cyclinDl表达下调,细胞增殖停滞。
5.FAK与细胞凋亡:Fujii等发现在原发性肝癌细胞中FAK的表达增加。30%的严重酒精性肝硬化者并发肝癌,如合并HBV和(或)HCV感染,发生肝癌的可能性更大。FAK持续活化可能是酒精性肝病导致肝癌的因素之一。
失巢凋亡是指正常细胞在失去ECM、缺少黏附的情况下发生的细胞凋亡,这一机制能防止失去ECM联系的细胞在不适宜的位置黏附生长。过度表达FAK可能使细胞在缺乏黏附的情况下,转为非锚定生长,失去生长抑制而不断增生。应用小干扰RNA阻断FAK表达可明显增加细胞凋亡的水平,直接证明FAK参与抑制细胞的凋亡。
FAK-PI3K是一个非常重要的锚定依赖的生存信号,FAK能激活PI3K介导的蛋白激酶B/Akt凋亡信号途径,活化抗凋亡蛋白Bcl-2,使细胞免于凋亡。FAK通过转录因子Jun的磷酸化激活JNK/SAPK途径。ERK信号通路影响cyclinDl的表达,从而调节细胞周期。因此FAK过度表达抑制细胞凋亡。
FAK在酒精性肝病发病机制中的作用尚不完全清楚,可能与以下几个方面有关:慢性乙醇**可以上调黏着斑蛋白基因表达,增加整合素的聚集,而使FAK磷酸化水平上升,从而影响一系列下游的信号传导通路(图2)。FAK可通过各种通路激活ERK信号转导途径,增强HSL活性,激活转录因子Egr-l,促进KCTNFa表达,导致脂肪肝的发生。FAK诱导cyclinDl转录,促进HSC增殖,从而引起肝纤维化。另外FAK过度表达可抑制细胞凋亡,这可能与晚期肝癌的发生有关。
目前的研究只是证实在乙醇**的大鼠黏着斑蛋白基因表达上调,但FAK蛋白表达并没有显著改变,也没有直接证据证明FAK的磷酸化水平是否有变化,起主要作用的磷酸化位点、该位点激活后引起的下游效应、FAK-ERK通路在酒精性肝病发病机制中发挥的作用均待进一步研究。
本站所注明来源为"爱爱医"的文章,版权归作者与本站共同所有,非经授权不得转载。
本站所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们
联系zlzs@120.net,我们将立即进行删除处理
- 相关阅读
- · 肝硬化病因和临床表现
- · 结节性肝硬化的有效预防
- · 引起结节性肝硬化的病因
- · 结节性肝硬化有什么症状表现
- · 2016年中西医医师考试综合笔试:肝硬化的治疗
热点图文
-
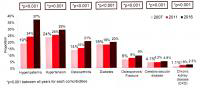
AASLD2018研究进展丨慢乙肝患者治疗期间的肾脏安全管理
在新药取得成功之前,应用口服核苷(酸)类似物(NA)治疗慢性乙型肝炎(简称...[详细]
-
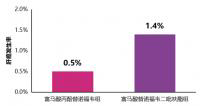
快讯丨TAF治疗4年的肝细胞癌发生率低于TDF
5月17日,在第十届全国疑难及重症肝病大会上,我国香港大学司徒伟基教授交流...[详细]
